Taeniodonts and
tillodonts:
Strange rooters and
diggers
The Early Cenozoic is well known for its archaic, yet
sometimes bizarrely adapted groups of mammals, which often lack
any direct living counterparts. Prime examples for this are the
taeniodonts and the tillodonts, two extinct orders of mammals
that evolved into a roughly similar direction: Both were
medium-sized to large animals that developed their front teeth
into great tusks and had claws at their feet (1,
2, 3). Once regarded as
closely related, it is now clear that both groups had a
completely separate history.
The order Taeniodonta is a
thoroughly North American group. Although fossil finds of
taeniodonts are widely distributed across North America, these
animals are rare in all known fossil assemblages. The first
taeniodonts appear very early in the Paleocene, perhaps not more
than 500000 years after the end of the Cretaceous. They probably
evolved from insectivore-like
animals such as Cimolestes and Procerberus that
occur in the Latest Cretaceous and Early Paleocene of the Rocky
Mountain region. Possible transitional forms have been reported
from the Earliest Paleocene of the same area but still remain to
be described in detail (1, 4).
 The most primitive taeniodonts were
still quite generalized animals. Onychodectes from the
early Paleocene of New Mexico is their best known representative
(1, 4). With the size of a
large house cat or a small dog, Onychodectes was
significantly larger than its insectivorous ancestors, yet its
skeleton is still comparable to primitive mammals like the
opossum. The skull of Onychodectes was long and narrow,
quite insectivore-like and very different from the shortened,
wide skulls of advanced taeniodonts. The canines that would later
become great tusks were still moderate in size. Onychodectes
had robust limbs, in particular a powerful forearm, and a mobile
hand, which suggests a certain propensity for digging. Hands and
feet each carried five digits with claws, and the tail was
extremely long and heavy. It is reasonable to suspect that Onychodectes
was a good climber, and the tail may have been prehensile to a
certain degree. According to its dentition the animal was an
omnivore. The sharp incisors and canines and the cutting
premolars that Onychodectes had inherited from its
ancestors were probably utilized to feed on insects, eggs and
small vertebrates. As the well developed grinding capabilities of
its posterior cheek teeth show, the animal also took a large
amount of plant matter. Onychodectes may have used its
relatively heavy forelimbs and the claws to dig up vegetables
hidden in the ground. The large amount of grit that is inevitably
mixed with such food would explain the heavy abrasion which can
be observed on teeth of Onychodectes.
The most primitive taeniodonts were
still quite generalized animals. Onychodectes from the
early Paleocene of New Mexico is their best known representative
(1, 4). With the size of a
large house cat or a small dog, Onychodectes was
significantly larger than its insectivorous ancestors, yet its
skeleton is still comparable to primitive mammals like the
opossum. The skull of Onychodectes was long and narrow,
quite insectivore-like and very different from the shortened,
wide skulls of advanced taeniodonts. The canines that would later
become great tusks were still moderate in size. Onychodectes
had robust limbs, in particular a powerful forearm, and a mobile
hand, which suggests a certain propensity for digging. Hands and
feet each carried five digits with claws, and the tail was
extremely long and heavy. It is reasonable to suspect that Onychodectes
was a good climber, and the tail may have been prehensile to a
certain degree. According to its dentition the animal was an
omnivore. The sharp incisors and canines and the cutting
premolars that Onychodectes had inherited from its
ancestors were probably utilized to feed on insects, eggs and
small vertebrates. As the well developed grinding capabilities of
its posterior cheek teeth show, the animal also took a large
amount of plant matter. Onychodectes may have used its
relatively heavy forelimbs and the claws to dig up vegetables
hidden in the ground. The large amount of grit that is inevitably
mixed with such food would explain the heavy abrasion which can
be observed on teeth of Onychodectes.
Figure 1: The primitive taeniodont Onychodectes
climbing up a tree in pursuit of food. Skull and body length
about 60 cm (4). From
(1).
Generalized taeniodonts like Onychodectes only enjoyed
a short-lived success, persisting no longer than until the Middle
Paleocene. By this time better adapted herbivores and predators
were narrowing the ecospace that had previously been available to
such unspecialized animals. More progressive taeniodonts had
already firmly occupied the niche of specialized rooters and
diggers and were thus preventing their generalized cousins from
evolving successfully into this direction (1, 4).
The more progressive taeniodonts, or stylinodontids, were in
fact among the animals that evolved most rapidly after the
disappearance of the dinosaurs - a classical example of so-called
quantum evolution. While their Cretaceous ancestors did not
exceed rat size, the Early Paleocene stylinodontid Wortmania
already attained a body mass of about 20 kg, perhaps as little as
500000 years after the K/T boundary. All important adaptations of
the stylinodontids are at least foreshadowed in Wortmania.
About two million years later, the stylinodont body plan was
essentially established with Psittacotherium, a robust
animal of about 50 kg. Wortmania and Psittacotherium
were each among the largest mammals of their time. Later
stylinodontids only grew little in size. Ectoganus of the
Late Paleocene to Early Eocene includes species both larger and
smaller than Psittacotherium. The last of the taeniodonts,
Early to Middle Eocene Stylinodon, reached a body mass of
up to 80 kg and was roughly pig-sized, though there was also a
rare dwarfed variant. Several much larger animals had evolved by
this time, including the gigantic uintatheres (1, 4, 5).
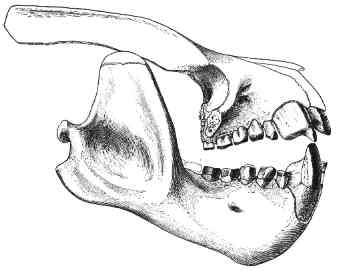
Figure 2: Partly preserved skull of the Middle to Late
Paleocene taeniodont Psittacotherium,
showing huge canines and massive jaws. Total skull length about
25 cm (4). From (13).
Psittacotherium, the "parrot beast",
exemplifies well the striking specializations that the
stylinodontids evolved for food processing. Its skull was heavily
built, with a shortened face and very robust jaws, trends that
were further elaborated in later stylinodontids. As areas of
muscle attachment show, the jaw musculature was powerful and the
tongue was large and well-developed. In the dentition emphasis
was clearly on the front teeth, which formed an impressive
cutting apparatus. The canines were greatly enlarged, as were the
upper incisors, though to lesser extent. Like in the gnawing
teeth of rodents the hard enamel was limited to the front of
these teeth, while the softer dentine was exposed at the rear,
which ensured that the chisel-like teeth always had sharp cutting
edges. The surface on the back served for crushing and grinding.
In Ectoganus and Stylinodon the canines became
ever-growing. The cheek teeth of these advanced stylinodontids
were reduced to simple grinding pegs. They lost their enamel
covering early in the life of an individual because of heavy
wear. The full set of teeth was ever-growing in Stylinodon
(1, 4).
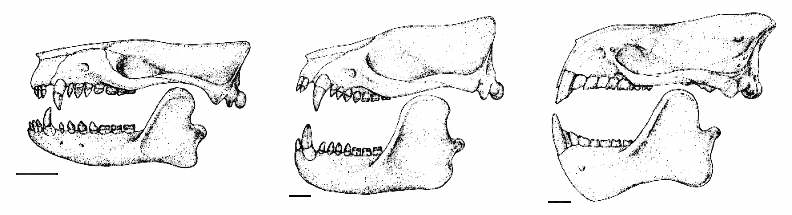
Figure 3: Reconstructed skulls of Early Paleocene Onychodectes
(left) and Wortmania (middle) and Late
Paleocene to Early Eocene Ectoganus (right),
scale bar 2 cm. Although not necessarily forming a single
ancestor-descendant lineage, these taeniodonts illustrate how
inconspicuous insectivore-like animals evolved into some of the
most bizarre creatures of the Early Cenozoic. After (4).
As their skeleton testifies, the stylinodontids were heavily
built animals. Their neck was short and stout. The limbs were
robust and powerful, especially the forelimbs with their
prominent crests and processes for the attachment of muscles.
Both hands and feet had five digits, the first and fifth one much
reduced on the forelimb, all equipped with large claws. The claws
of the hand are particularly striking in being recurved and
laterally compressed - a feature already present in the earliest
stylinodontid Wortmania. Together with the powerful
musculature of the forelimb, these claws must have rendered the
stylinodontids excellent diggers. On the foot the claws are
broader, and the digits are arranged in an arc. They provided
firm support during digging, together with the long and heavy
tail (1, 4).
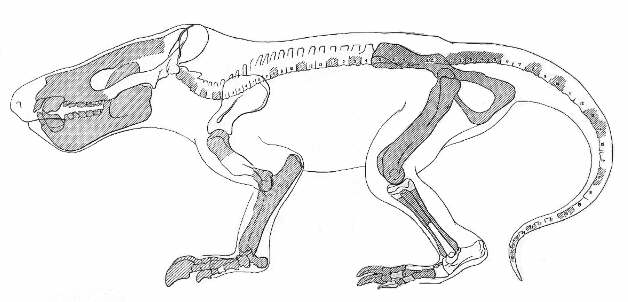
Figure 4: Skeleton of the taeniodont Psittacotherium.
Though this restoration from the 1930s is not
completely up to date, it still illustrates well the digging
adaptations of the forelimb, including large claws and a
prominent posterior process of the ulna. Strong muscles attached
to the latter for the retraction of the forelimb. Skull and body
length about 110 cm (4).
From (13).
In terms of their skeleton, advanced taeniodonts such as Ectoganus
or Stylinodon can be compared to the aardvark Orycteropus,
an active digger. Like this recent mammal, the stylinodontids may
have dug burrows to live in. Yet their powerful masticatory
apparatus indicates that they fed on more resistant things than
the ants and termites which the aardvark prefers, presumably some
kind of tough plant matter that was ripped and torn with claws
and teeth. Using their powerful forelimbs and claws, the
stylinodontids may have dug for tubers, roots and other
underground vegetables. Like toady's pigs and peccaries, they may
also have utilized their powerful snouts and enlarged canines for
digging and rooting. In analogy to these recent mammals, it seems
reasonable that stylinodontids did not refrain from eating
carrion or other animal matter if they happened to stray across
it (1, 4).
The stylinodontids seem to have been relatively slow-moving
animals, adapted for strength at the expense of speed. Judging
from their small brain size, they were not particularly bright.
While the stylinodontids of the Eocene may have been hunted by
advanced carnivorous mammals (4), earlier
forms like Psittacotherium were probably not seriously
threatened by the primitive predators
of their time thanks to their large size, except at the waterside
where crocodiles lurked. Even encounters with these dangerous
reptiles may have been rare, since it has been suggested that the
taeniodonts were mainly open-country and upland forms (1, 4). Roots and tubers
contain large amounts of water and nutrients, and access to these
natural storages may have enabled the taeniodonts to invade drier
regions than it was possible for animals depending on vegetation
above the ground. If this hypothesis is correct, taeniodonts
could be rare in the fossil record simply because most Early
Tertiary deposits in North America represent animal communities
close to rivers. In their preferred upland habitat, taeniodonts
could have been much more common than the fossil record reflects.
In analogy to the aardvark, we may imagine them as solitary
animals, combing the open landscape in search of food and
returning to their burrows for shelter at the end of their daily
trips.

Figure 5: Restoration of the advanced taeniodont Ectoganus,
together with two large herbivores of the genus Pantolambda
that belong to another extinct order of mammals, the Pantodonta.
The scene is from the Middle Paleocene of the Rockies according
to (17), but in fact Ectoganus
was somewhat later in age.
Members of the second early Tertiary
order of large herbivorous mammals with claws, the Tillodontia,
were in many ways similar to the taeniodonts (2,
4, 6). Like the latter,
tillodonts evolved a pair of enlarged, chisel-shaped front teeth.
This was once interpreted as evidence of close relationships
between taeniodonts and tillodonts. However, the rodent-like
teeth represent different tooth positions in the two groups (the
canines in taeniodonts and the second incisors in tillodonts) and
must therefore have evolved independently. Although this had
already been recognized by the end of the nineteenth century,
finding more convincing links between tillodonts and other groups
of mammals proved to be a difficult task.
Identifying relatives of the tillodonts was long hindered by
the fact that the only substantial remains of the group came from
North America. Typical tillodonts appear abruptly on this
continent with Esthonyx (2, 6, 7), a genus ranging from
the latest Paleocene to the Middle Eocene, although the early
species are sometimes referred to a separate genus Azygonyx
(8). Esthonyx already foreshadows the
chisel-like second incisors of the advanced tillodonts, with the
enamel becoming restricted to the front side of the teeth, but
the canines and the third upper incisor are large as well in this
genus. Later tillodonts like Trogosus and Tillodon
further emphasized their second incisors, which became
ever-growing, while other teeth in the front of the jaw were
either reduced or lost (2, 6).
Together with a lengthening of the snout this led to the
development of gaps in the jaws between the enlarged tusks and
the cheek tooth battery - an arrangement vaguely similar to that
in rodents but very different from that in the short-snouted
taeniodonts. Like the latter group, the tillodonts disposed of
powerful jaw muscles, which were already well-developed in Esthonyx,
and their teeth are often heavily worn. This suggests that the
animals ingested a lot of soil and sand with their food, which
may have consisted largely of roots and tubers. Grooved wear
patterns on the enlarged incisors of Esthonyx provide
further clues to the feeding habits of the tillodonts. These
marks may have been produced as the animal was pulling roots out
of the earth or stripping off leaves or buds with its tusks (2, 7).
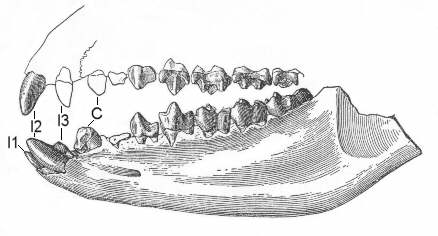
Figure 6: Dentition of the Latest Paleocene to Early Eocene
tillodont Esthonyx. Incisors (I1, I2, I3)
and canines (C) are labeled in the drawing. As is typical of
tillodonts, the chisel-like second incisors are particularly
well-developed, although the third upper incisor and the canines
are still prominent, too. Preserved jaw length about 10 cm. After
(6).
The first North American tillodonts were still medium-sized
animals, with body masses of approximately 10 to 30 kg in the
different species of Esthonyx (8).
Later members of the order, like the Early to Middle Eocene Trogosus,
became much larger, weighing about 150 kg or more. These
tillodonts were stout, compact animals that may have been almost
bear-like in appearance. Their hands bore moderately large and
recurved claws, and the musculature of their forelimb was
powerful. This may indicate well developed digging capabilities
like in taeniodonts, though the evidence is less conclusive than
in the latter case. Perhaps the advanced tillodonts used a
combination of digging and tearing with their claws and pulling
with their front teeth to obtain food (2, 3). Our knowledge of the anatomy of Esthonyx
is still very limited, despite the fact that this genus was
moderately common in the latest Paleocene to Early Eocene of the
Western Interior. What is known of its skeleton suggests rather
generalized capabilities of locomotion. Like in later tillodonts,
the feet of Esthonyx were equipped with laterally
compressed, recurved claws which were perhaps also utilized for
climbing (8, 9, 10). Again, the very small brain of Esthonyx
suggests that these archaic mammals were not particularly
intelligent (7).
Apart from Esthonyx, the only pre-Eocene mammal from
North America that could possibly be a tillodont is the enigmatic
Deltatherium from the Middle Paleocene of New Mexico (11, 12) (see here for life restoration). Although the
cheek teeth of Deltatherium somehow approach those of the
tillodonts, its anterior dentition looks very different, with
small incisors but large saber-like upper canines that almost
foreshadow those of the saber-toothed cats (13).
If actually related to the tillodonts, Deltatherium must
represent an early offshoot that branched away into a different,
more carnivorous direction and was not involved in the ancestry
of the typical tillodonts.
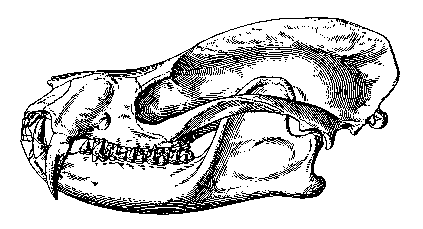
Figure 7: Skull of Deltatherium fundaminis,
a possible Middle Paleocene relative of the tillodonts that
evolved saber-like upper canines. Skull length about 14 cm. From
(12).
The sudden appearance of Esthonyx in the Latest
Paleocene of North America thus apparently represents an
immigration event, just like the incursion of tillodonts into
Europe at the beginning of the Eocene (this time probably from
North America, which was then connected to Europe via Greenland).
During the last decades, discoveries of primitive tillodonts in
the Paleocene of South China have made it possible to identify
Asia as the probable place of origin of the order (2,
14, 15). Among the Chinese
forms, Meiostylodon is closest to the first North American
tillodonts of the genus Esthonyx. The Chinese genus is
still smaller in size, yet it definitely shows the characteristic
chisel-shaped incisors of the group. Meiostylodon was
found in sediments from the earlier part of the Paleocene and is
therefore much older than Esthonyx. It may be closely
related to the ancestor of the latter which migrated into the New
World across a land bridge in the region of today's Bering
Strait, probably as climatic conditions became milder in these
high latitudes at the close of the Paleocene. Another probable
tillodont from China, Interogale, comes from late
Paleocene rocks but is nevertheless the smallest known member of
the order, about half the size of small species of Esthonyx.
Finally, an extremely primitive tillodont-like form, called Lofochaius,
is known from the earlier Paleocene of China. Lofochaius
may be close to the base of the tillodont radiation if it
represents in fact a member of this order. These and other
enigmatic forms from China are still poorly known, and more
fossils from this region are required to get a clearer picture of
the early evolution of the Tillodontia.
The Chinese discoveries also stimulated new discussions on the
relationships between tillodonts and other mammals. Based in part
on the new finds, the tillodonts are now usually regarded as the
sister group of another now extinct order of herbivorous mammals
that arose in Asia, the pantodonts
(2, 11) (see also restoration above).
Some advocates of this idea have even gone so far to include the
tillodonts in the order Pantodonta (16). In
contrast to earlier ideas, tillodonts and pantodonts do not seem
to belong to the hoofed mammals or Ungulata,
a group to which most large placental herbivores tended to be
assigned in the past. Instead, they may be direct descendants of
primitive insectivore-like mammals, just like the taeniodonts.
Both taeniodonts and tillodonts are part of the first wave of
archaic mammals which dominated the Paleocene faunas but
succumbed to competition with more modern mammals in the Eocene.
Advanced members of the Taeniodonta and Tillodontia were probably
ecologically similar to a certain degree, so they may also have
competed with each other in North America where both groups
occurred. This may have contributed to the disappearance of the
tillodonts from this continent in the early Middle Eocene, while
the order persisted into the Late Eocene in Asia. The last
taeniodonts of the genus Stylinodon managed to survive
until the late Middle Eocene in North America. By this time, they
had to face increasing competition with the even-toed ungulates
or Artiodactyla, in particular with pig-like forms that probably
had similar digging and rooting habits but were larger-brained.
These modern hoofed mammals were apparently capable of utilizing
similar resources in a more efficient way. Though the impressive
food processing apparatus of the taeniodonts had served its
purpose well for a long time in the absence of more efficient
competitors, it still seems to have been a rather crude solution,
and highly wasteful in energy. Extremely specialized but poor in
diversity as the last taeniodonts were, they were unable to
produce new adaptive patterns as the old ones proved to be no
longer successful. They were trapped in their narrow ecological
niche which they had occupied for more than 20 million years but
were finally unable to maintain (1, 2, 4).
<< Previous
article
>> Next
article
Back to introduction
First upload 01.11.02.
References:
- Lucas, S. G., Schoch, R.
M. & Williamson, T. E. 1998: Taeniodonta. In: Janis,
C. M. (ed.): Evolution of Tertiary Mammals of North
America. Cambridge University Press, 260-267.
- Lucas, S. G. &
Schoch, R. M. 1998: Tillodontia. In: Janis, C. M. (ed.):
Evolution of Tertiary Mammals of North America. Cambridge
University Press, 268- 273.
- Coombs, M. C. 1983:
Large Mammalian Clawed Herbivores: A Comparative Study.
Transactions of the American Philosophical Society 73,
1-96.
- Schoch, R. M. 1986:
Systematics, Functional Morphology and Macroevolution of
the Extinct Mammalian Order Taeniodonta. Yale University
Peabody Museum of Natural History Bulletin 42, 1-307.
- Sloan, R. E. 1987:
Paleocene and latest Cretaceous mammal ages, biozones,
magnetozones, rates of sedimentation, and evolution.
Geological Society of America Special Paper 209, 165-200.
- Gazin, C. L. 1953: The
Tillodontia: an Early Tertiary order of mammals.
Smithsonian Miscellaneous Collections 121, no. 10, 1-110.
- Gingerich, P. D. &
Gunnell, G. F. 1979: Systematics and evolution of the
genus Esthonyx (Mammalia, Tillodontia) in the
early Eocene of North America. Contributions from the
museum of Paleontology, University of Michigan, 25,
125-153.
- Gingerich, P. D. 1989:
New earliest Wasatchian mammalian fauna from the Eocene
of northwestern Wyoming: composition and diversity in a
rarely sampled high-floodplain assemblage. University of
Michigan Papers on Paleontology 28, 1-97.
- Rose, K. D. 1990:
Postcranial remains and adaptations in early Eocene
mammals from the Willwood Formation, Bighorn Basin,
Wyoming. In: Bown, T. M. & Rose, K. D. (eds.): Dawn
of the Age of Mammals in the Northern Part of the Rocky
Mountain Interior. Geological Society of America Special
Paper 243, 107-133.
- Rose, K. D. 2001:
Compendium of Wasatchian Mammal Postcrania from the
Willwood Formation of the Bighorn Basin. In: Gingerich,
P. D. (ed.): Paleocene-Eocene Stratigraphy and Biotic
Change in the Bighorn and Clarks Fork Basins, Wyoming.
University of Michigan Papers on Paleontology 33,
157-183.
- Lucas, S. 1993:
Pantodonts, Tillodonts, Uintatheres, and Pyrotheres Are
Not Ungulates. In: Szalay, F. S., Novaceck, M. J. &
McKenna, M. C. (eds.): Mammal Phylogeny. Volume 2:
Placentals, 182-194.
- Williamson, T. E. 1996:
The Beginning of the Age of Mammals in the San Juan
Basin, New Mexico: Biostratigraphy and Evolution of
Paleocene Mammals of the Nacimiento Formation. New Mexico
Museum of Natural History and Science Bulletin 8, 1-141.
- Matthew, W. D. 1937:
Paleocene faunas of the San Juan Basin, New Mexico.
Transactions of the American Philosophical Society, new
series, 30, 1-510.
- Ting, S. & Zheng, J.
1989: The affinities of Interogale and Anchilestes
and the origin of the Tillodontia. Vertebrata PalAsiatica
27, 77-86.
- Wang, Y., Hu, Y., Chow,
M. & Li, C. 1998: Chinese Paleocene mammal faunas and
their correlation. In: Beard, K. C. & Dawson, M. R.
(eds.): Dawn of the Age of Mammals in Asia. Bulletin of
Carnegie Museum of Natural History 34, 89-123.
- Chow, M. & Wang, B.
1979: Relationships between pantodonts and tillodonts and
classification of the order Pantodonta. Vertebrata
Palasiatica 7, 97-104. (Chinese with English abstract)
- Savage, R. J. G. &
Long, M. R. 1986: Mammal evolution. An illustrated guide.
British Museum (Natural History).