Predation on mammals, among others by dinosaurs, was without doubt an important factor in the early evolution of this class of animals. There are, on the other hand, only few Mesozoic mammals that could have been carnivorous, and probably none preyed on larger vertebrates.
After the dramatic turnover at the K/T boundary, the mammals were facing a completely different situation. In the earliest Paleocene there is only very limited evidence of terrestrial carnivores, and it has even been claimed that the effect of predation on mammals was negligible. Terrestrial crocodiles, snakes and larger lizards survived the K/T boundary and are potential predators, and aquatic crocodiles certainly also took the opportunity to catch unwary animals drinking at the river banks. Nevertheless, pressure from reptiles was clearly much reduced on land. Large flightless birds have been found in later Paleocene rocks in some parts of the world, like Gastornis in Europe, and could have hunted mammals. However, they are entirely unknown from the rich fossil record of the North American Paleocene: The famous Diatryma first appears there in the early Eocene. Knowledge of other birds is very limited. The earliest representatives of the owls, which typically hunt mammals, are Ogygoptynx and Berruornis from the late Paleocene of the U. S.A. and France respectively. Characteristic nodules containing remains of small mammals occur at a late Paleocene locality in China. They have been interpreted as pellets regurgitated by birds and form the first direct evidence of birds preying on mammals.
On the other hand, the mammals of the earliest Paleocene were nearly all rat-sized, and their teeth do not yet show clear specializations for cutting meat. Animal communities on land were thus characterized by a uniquely low amount of predation just after the K/T turnover. The step towards the first meat-eating mammals is, however, not such a dramatic one. Primitive mammals typically fed on insects, and there is a smooth transition between an insectivore that occasionally takes small vertebrates and a small predator. In South America this led to the evolution of carnivorous marsupials like the borhyaenids. Two major groups of placental mammals made this transition during the Paleocene, the now extinct creodonts (order Creodonta) and the true carnivores (order Carnivora) that flourish today. Possible ancestors for both of them are the shrew-like paleoryctids from the late Cretaceous to early Tertiary, so the two groups could be closely related. Both creodonts and true carnivores differ from these primitive insectivores in having special pairs of upper and lower cheek teeth called carnassials that cut meat like a pair of scissors. As the jaws close, a blade at the rear of the upper carnassial shears past the sharp front edge of the following lower carnassial. Members of the Carnivora have only one pair of carnassials, which always consists of the last upper premolar and the first lower molar. Creodonts often have more than one pair of carnassials, which are always formed by the molars. Members of both orders have large canines for killing their prey.
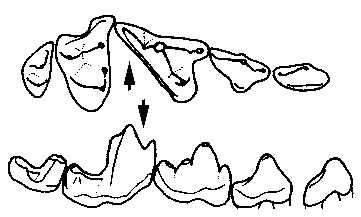
Figure 1: Cheek teeth of the true carnivore Protictis from the Paleocene of North America. Upper teeth above, looking at the chewing faces. Lower teeth below, seen in profile from outside. Front of the jaw is to the right. The blades of the enlarged carnassials are marked with arrows. Modified after Flynn, in Janis et al. (eds.) (1998).
True carnivores of the order Carnivora appear first in the middle or even early Paleocene of North America. They form an important component of many Paleocene faunas of this continent, which may be their place of origin. Members of the Carnivora remain small, generalized predators throughout the Paleocene and during much of the Eocene, comparable in ecology to the martens and civets of today. Most of the Paleocene forms belong to the family Viverravidae, a primitive group with only one major specialization, the loss of their last molars. Since cats and their relatives have also lost their last molars, the viverravids are sometimes regarded as ancestors of this group within the Carnivora. Protictis (meaning "pre-weasel") is a typical viverravid known from several Paleocene species. Although the characteristic carnassials are clearly developed, the high, sharp teeth of Protictis show that insects still made up an important part of its diet. A fossil brain cast suggests that both vision and hearing were important senses for this early predator. However, the position of the orbits in early carnivores indicates that three-dimensional vision was still much less developed than in modern carnivores. The known skeletal remains of Protictis suggest that it was living on the forest floor. A second family of small generalized carnivores, the Miacidae, appears with Uintacyon in the latest Paleocene. This group flourished in the early Eocene and included forms that were equally adapted to life in the trees and on the ground.

The generally larger creodonts are first known from later Paleocene time. They soon became the dominant carnivorous mammals of the early Tertiary all over the world, with the exception of South America, Australia and probably Antarctica. The first definitive creodonts are members of the family Oxyaenidae from the late Paleocene of North America. Oxyaenids have been compared to martens or cats among todays carnivores. They were animals with a long body, short limbs and and a very long tail. Oxyaenids were walking more or less on flat feet, not on their toes like later carnivores adapted to running. The large cat-like genus Oxyaena is best known from the early Eocene. Related late Paleocene species are smaller (estimated body weight 3 to 8 kg) and have less well developed carnassials. They are classified as Oxyaena by some authors but as a different genus Dipsalidictis by others. Whatever the classification, these animals were probably generalized carnivores that fed on birds, small mammals, eggs and perhaps still some insects, similar to the living civets. The mobile structure of their feet suggests that they were capable of climbing in trees. The late Paleocene Dipsalodon and the late Paleocene to early Eocene Palaeonictis represent a different adaptive line of oxyaenids. Their much heavier jaws and blunt, robust teeth were more suited for crushing bones than for slicing meat. This adaptation is typically found in scavenging animals. The biggest species was Palaeonictis peloria (meaning "terrible ancient weasel") from the latest Paleocene of Wyoming, U.S.A. It is known from an incomplete jaw that must have measured well over 20 cm in length. This animal was the largest carnivore in its ecosystem.
Figure 2: Reconstruction of the cat-like creodont Oxyaena, based on early Eocene species. Late Paleocene species of Oxyaena or Dipsalidictis were probably similar but smaller. From Benes & Burian (1980).
A second branch of creodonts were the highly successful hyaenodontids, typically dog- or hyaena-like predators with improved capability of running. For a long time they have only been known from Eocene and later rocks, except for some questionable specimens. The sudden appearance of hyaenodontid creodonts at the beginning of the Eocene (together with several other groups of mammals) has been interpreted as an immigration from some part of the world with a poor fossil record for the Paleocene. This place could have been Africa, a stronghold of hyaenodontids in later time, but positive evidence is still missing. The oldest unquestionable fossils of hyaenodontid creodonts have recently been discovered in the latest Paleocene of the Mongolian Plateau. They include a primitive species of Prolimnocyon, a small generalized predator already known from the early Eocene.
As we have seen, neither carnivores nor creodonts evolved larger meat-eating forms before the late Paleocene. Yet this ecological niche had not been completely vacant before. It had been partly occupied by a very different group which we would hardly associate with carnivorous habits today: The ungulates or hoofed mammals.
Primitive ungulates are united in the somewhat artificial order Condylarthra. The condylarths form the stem group from which probably all hoofed mammals arose. The earliest condylarths were small omnivores that may have fed on soft plants, fruits and insects. Many of their descendants became more herbivorous, but at least one group of condylarths, the arctocyonids, sticked to an omnivorous diet and even acquired carnivorous habits to some extent. Although arctocyonids never were specialized predators, it is probable that they increasingly included animal prey in their diet as they grew in size. By middle Paleocene time arctocyonids had reached the size of a wolf or a small bear, like Arctocyon from Europe and Claenodon (a possible synonym of Arctocyon) from North America. Their anatomy is remarkably primitive and suggests that they were rather clumsy animals, with short, stout limbs and an extremely long, heavy tail. Though definitely not built for running down their prey, their powerful proportions, sharp claws and large canines may have enabled them to overpower smaller animals in a surprise attack. The cheek teeth of Arctocyon and Claenodon are broad and flat, very different from those of specialized carnivorous mammals but similar to those of bears. This indicates that, like these modern omnivores, Arctocyon and Claenodon still fed to a large degree on fruits, nuts, seeds and insects.
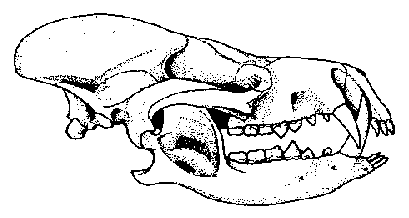
Figure 3: Skull of Arctocyon primaevus from the late Paleocene of France, a condylarth that probably included meat in its diet. Note the large canines and the low bear-like cheek teeth at the back of the jaw. Skull length about 25 cm. From Russell (1964).
It is noteworthy that some arctocyonids started to develop a saber-tooth dentition, the type of teeth that is most widely known from saber-toothed cats but occurs independently in other mammals like the marsupial saber-tooth Thylacosmilus. Among arctocyonids this trend can first be seen in Mentoclaenodon from the middle Paleocene fissure fillings of Walbeck, Germany. With an estimated skull length of about 15 cm this rare form is the largest mammal of the Walbeck fauna. Mentoclaenodon has particularly large upper canines that are crenulated at the back, and it shows a beginning expansion of the chin region as this is often associated with saber-teeth. In Anacodon, the late Paleocene to early Eocene terminal form of the large North American arctocyonids, the chin is developed into a large flange to protect the saber-like upper canines. These arctocyonids may have been the first carnivore-like mammals ever to evolve this type of dentition.
A closely related group of ungulates, the order Mesonychia, advanced more strongly into the direction of carnivores than the arctocyonids. Paleocene members belong mainly to the families Triisodontidae and Mesonychidae. The triisodonts were an early offshot consisting of animals with massive jaws and heavily built cheek teeth. In North America they were by far the largest mammals in the early Paleocene: Just after the K/T boundary, when all other mammals hardly exceeded rat size, the triisodontid Eoconodon was nearly an order of magnitude larger in body weight. Its middle Paleocene descendant Triisodon surpassed any arctocyonid in size. Together with the arctocyonids the triisodontids may have partly replaced true carnivorous forms in the early mammal communities. Interestingly, Andrewsarchus from the middle Eocene of Asia has been regarded as a gigantic (skull length 84 cm) late survivor of the triisodontid line. This genus has been called the largest carnivorous mammal of all times, but it was probably rather an omnivore.
Of all ungulates, the mesonychids included probably the most dedicated terrestial predators. They can be compared to dogs, hyaenas and bears in general appearance. Their main weapons for attacking prey were the large, pointed canines, a typical trait of carnivorous mammals. Unlike other predators, however, mesonychids did not dispose of sharp claws to tear their prey. Instead, their feet retained a telltale sign of their ungulate ancestors, which is quite unusual for a carnivore: Hoofs. The cheek teeth of mesonychids were modified for cutting meat, but the shearing mechanism was very different from - and obviously less effective than - the carnassial structure that carnivores, creodonts and carnivorous marsupials acquired. In fact, mesonychids may have used their teeth mainly for grasping chunks of meat, which were torn off with movements of the head while the feet anchored the prey.
Mesonychids probably originated in Asia, where the most primitive mesonychid, Yangtanlestes, is known from the earlier part of the Paleocene. They were also most diverse in Asia, where they occur in all major Paleocene faunas. Since true carnivores, creodonts and condylarths were either rare or completely absent in these animal communities, mesonychids obviously dominated the large predator niche in the Paleocene of Asia. Two genera of mesonychids, Dissacus and Ankalagon, succesfully spread to other continents in the middle Paleocene. Dissacus was a coyote-sized carnivore that has been found all over the northern hemisphere. Ankalagon from the middle Paleocene of New Mexico, U. S. A. (reconstructed here in its natural environment) was much larger, with a skull comparable in size to that of a brown bear. However, like many archaic animals, mesonychids had a relatively large head, so the body of Ankalagon was smaller than that of the modern carnivore. Early mesonychids still had five digits on their feet, which probably rested flat on the ground during walking. Later mesonychids were increasingly well adapted to running. An example is the latest Paleocene to Eocene Pachyaena, a wolf- to bear-sized mesonychid. Like running members of the even-toed ungulates, Pachyaena walked on its digits, which were reduced to four in number and ended in hoofs. Its anatomy suggest that this mesonychid was adapted more for endurance than for speed in running. Smaller species of Pachyaena may have been more carnivorous, larger species more omnivorous in habits. The dentition of Pachyaena shows no special signs of bone-crushing, unlike the teeth of some later mesonychids that specialized on carrion-feeding.

Figure 4: Flighless birds of the genus Diatryma (about 2.4 m in height) try to defend their chicks against a pack of Pachyaena. Although the mesonychid Pachyaena is known from the latest Paleocene, this encounter must be placed in the early Eocene when Diatryma first appears. Smaller mesonychids of the genus Dissacus occur together with the large flightless bird Gastornis in the Paleocene of Europe. Drawing by Gregory S. Paul, from Stanley (1989).
The transition from the Paleocene carnivore assemblage to the modern assemblage of true carnivores took place step by step and was not always synchronous around the world. Archaic forms like the arctocyonids and triisodontids disappeared soon after the Paleocene. The last mesonychids survived into the Oligocene in Asia. Their decline may be linked to competition with creodonts and true carnivores. The creodonts were the dominant predators of the Eocene. However, their diversity quickly declined after the appearance of more modern Carnivora in the late Eocene, and the last creodonts got extinct in the late Miocene. There have been speculations that the creodonts were inferior to the true carnivores in anatomy or intelligence, but their fossils supply no evidence for this. So it remains a mystery why this great radiation of advanced predators vanished and only one single group of placental meat-eaters remained on land, the Carnivora.
Yet one branch of archaic predators may have survived to this day in the oceans: The whales. Molecular biologists link them generally to the even-toed ungulates, but many paleontologists favor a descent from the Mesonychia. The most primitive whales from early Eocene coastal deposits in Asia show some striking similarities to mesonychians. So we may speculate that a fish-eating population of Paleocene mesonychians living on the shores of Asia returned to the sea, forming the beginning of a ratiation that led to the largest animals ever in existence.
[Discoveries of early whales published in September 2001 provide strong evidence that whales have their origin in even-toed ungulates. In this case, much convergent evolution must have happened between mesonychians and early whales.]
Major sources for this article:
Benes & Burian 1980: Tiere der Urzeit. Verlag Werner Dausien.
Carroll, R. L. 1988: Vertebrate paleontology and evolution. Freeman and Company.
Gingerich, P. D. & Winkler, D. A. 1985: Systematics of Paleocene Viverravidae (Mammalia, Carnivora) in the Bighorn Basin and Clark's Fork Basin, Wyoming. Contributions from the Museum of Paleontology, University of Michigan, 26, 197-225.
Gunnell, G. F. & Gingerich, P. D. 1991: Systematics and evolution of late Paleocene and early Eocene Oxyaenidae (Mammalia, Creodonta) in the Clarks Fork Basin, Wyoming. Contributions from the Museum of Paleontology, University of Michigan, 28, 141-180.
Janis, Ch. M., Scott, K. M. & Jacobs, L. L. 1998 (eds.): Evolution of Tertiary Mammals of North America. Volume 1: Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals. Cambridge University Press.
Matthew, W. D. 1937: Paleocene faunas of the San Juan Basin, New Mexico. Transactions of the Philosophical Society, v. 30, 1-510.
Meng, J., Zhai, R. & Wyss, A. 1998: The late Paleocene Bayan Ulan Fauna of Inner Mongolia, China. In: Beard, C. K. & Dawson, M. R. (eds.): Dawn of the Age of Mammals in Asia. Bulletin of Carnegie Museum of Natural History 34, 148 - 185.
O'Leary, M. A. & Rose, K. D. 1995: Postcranial skeleton of the early Eocene mesonychid Pachyaena. Journal of Vertebrate Paleontology 15, 401-430.
Russell, D. E. 1964: Les mammifères paléocènes d'Europe. Mémoires du Muséum National d'Histoire Naturelle, Série C, 8, 1-324.
Stanley, S. M. 1989: Earth and Life Through Time. Freeman and Company.
Szalay, F. S. & Gould, S. J. 1966: Asiatic Mesonychidae (Mammalia, Condylarthra). Bulletin of the American Museum of Natural History 132, 127-174.
Szalay, F. S. 1969: Origin and evolution of the mesonychid condylarth feeding mechanism. Evolution 23, 703-720.
Van Valen, L. 1978: The beginning of the Age of Mammals. Evolutionary Theory 4, 45-80.
Weigelt, J. 1960: Die Arctocyoniden von Walbeck. Freiberger Forschungshefte C.77, 1-241.